A cikk tartalma Show
A emberi test egy rendkívül komplex és finoman hangolt rendszer, ahol minden szervnek és sejtszerkezetnek pontosan meghatározott feladata van. Ezen bonyolult hálózat egyik legfontosabb, ám gyakran alulértékelt komponense a vese, amely létfontosságú szerepet játszik a szervezet belső környezetének, a homeosztázisnak a fenntartásában.
A vese nem csupán kiválasztószerv; a vér tisztítása mellett szabályozza a folyadék- és elektrolit-egyensúlyt, a vérnyomást, és hormonokat is termel. Ennek a sokrétű feladatnak a középpontjában a nefron áll, a vese mikroszkopikus funkcionális egysége.
A nefronon belül számos speciális szegmens található, amelyek mindegyike hozzájárul a vizeletképzés összetett folyamatához. Ezek közül az egyik legkiemelkedőbb és leginkább pótolhatatlan rész a Henle-kacs, amelynek egyedülálló anatómiája és fiziológiája alapvető a szervezet vízháztartásának és a koncentrált vizelet képzésének szempontjából.
Nélküle a szervezet képtelen lenne hatékonyan visszatartani a vizet, és a folyadékveszteség gyorsan életveszélyes dehidratációhoz vezetne. Ez a cikk a Henle-kacs mélyére ás, feltárva működésének bonyolult mechanizmusait és pótolhatatlan szerepét a vese szűrési folyamataiban, a molekuláris szinttől a klinikai jelentőségig.
A vese – Az élet alapköveinek őre
A vese páros szerv, amely a hasüreg hátsó részén, a gerincoszlop két oldalán helyezkedik el. Formája bab alakú, és mérete körülbelül egy ököl nagyságú. Bár viszonylag kis méretű, a szervezet teljes vérellátásának mintegy 20-25%-át kapja nyugalmi állapotban, ami rávilágít a feladatainak kritikus fontosságára.
Fő feladata a vér szűrése, a salakanyagok eltávolítása és a hasznos anyagok, mint például a víz és az elektrolitok visszatartása. Ezenkívül szerepet játszik a vérnyomás szabályozásában (renin termelésével), a vörösvértest-termelés stimulálásában (eritropoetin termelésével), és a D-vitamin aktiválásában is.
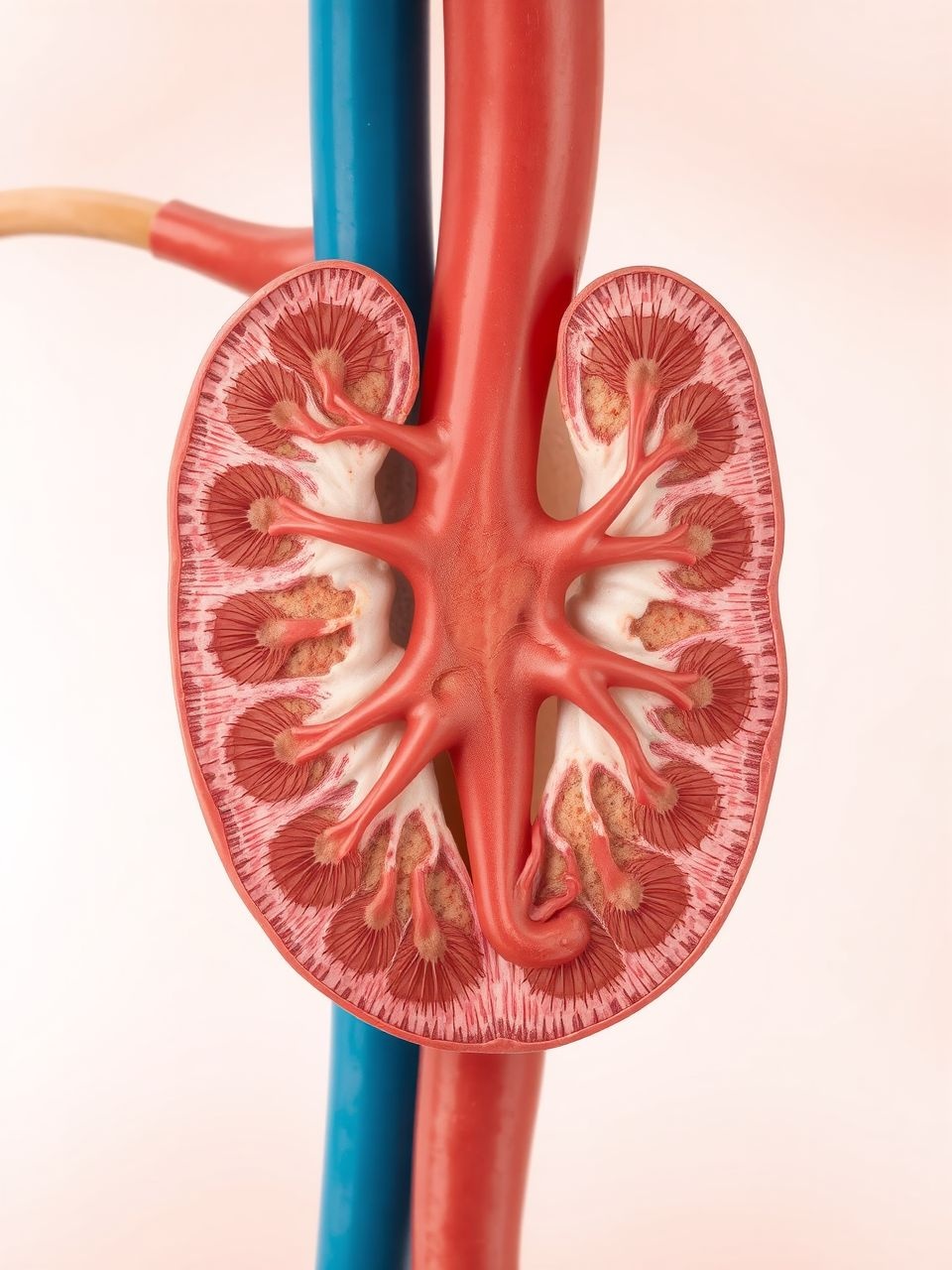
A vese belső szerkezete két fő régióra osztható: a külső kéregrészre (cortex) és a belső velőállományra (medulla). A velőállomány jellegzetes piramisokból áll, amelyek a vesemedencébe nyílnak. Ezek a makroszkopikus struktúrák szolgálnak otthonul a mikroszkopikus funkcionális egységeknek, a nefronoknak.
A vese működése tehát nem korlátozódik pusztán a vizelet kiválasztására. Egy komplex endokrin szervként is funkcionál, amely szorosan összehangolja a szervezet számos élettani folyamatát. A vesék egészsége elengedhetetlen a teljes test egészségének fenntartásához.
A nefron: A vese funkcionális egysége
Minden egyes vesében körülbelül egymillió nefron található, és ezek a mikroszkopikus egységek felelősek a vér szűrésének és a vizelet képzésének minden lépéséért. Egy nefron több, egymással szorosan összefüggő részből áll, amelyek mindegyike speciális feladatot lát el.
A vizeletképzés folyamata a glomerulusban kezdődik, amely egy kapillárisgombolyag a Bowman-tokban. Itt történik a vér ultrafiltrációja: a víz és a kis molekulatömegű oldott anyagok átszűrődnek a vérből a Bowman-tokba, miközben a nagyobb fehérjék és vérsejtek a vérben maradnak.
Az így keletkezett szűrlet, az úgynevezett primer vizelet, a proximális tubulusba (első elvezető csatorna) jut. Itt történik a szűrlet térfogatának legnagyobb részének visszaszívása, beleértve a glükózt, aminosavakat, sót és vizet. A proximális tubulus rendkívül aktív a transzportfolyamatokban.
Ezt követi a Henle-kacs, amelynek egyedülálló, U alakú szerkezete alapvető az ozmotikus gradiens kialakításában a vese velőállományában. Ez a gradiens teszi lehetővé a koncentrált vizelet képzését, ahogy azt részletesebben tárgyaljuk majd.
A Henle-kacs után a szűrlet a disztális tubulusba (második elvezető csatorna) kerül, ahol a finomhangolás történik. Itt az aldoszteron és az antidiuretikus hormon (ADH) hatására további só- és vízvisszaszívás, valamint kálium- és hidrogénion-szekréció zajlik, szabályozva az elektrolit-egyensúlyt és a pH-t.
Végül a szűrlet a gyűjtőcsatornákba áramlik, amelyek több nefronból gyűjtik össze a vizeletet. Itt az ADH hatására történik a végső vízvisszaszívás, ami lehetővé teszi a szervezet számára, hogy rendkívül koncentrált vizeletet ürítsen, ha szükséges. A gyűjtőcsatornák a vesemedencébe vezetik a már kész vizeletet.
A Henle-kacs anatómiája: Részletes áttekintés
A Henle-kacs a nefron azon része, amely a proximális és a disztális tubulus között helyezkedik el, és U alakú hurkot képez. Két fő típusa létezik a nefronoknak, amelyekben a Henle-kacs eltérő hosszúságú és elhelyezkedésű, de mindkettő alapvető a veseműködés szempontjából.
A kortikális nefronok (mintegy 85%) Henle-kacsa viszonylag rövid, és csak csekély mértékben nyúlik be a vese velőállományába. Fő feladatuk a szűrlet térfogatának csökkentése és az elektrolit-egyensúly fenntartása.
Ezzel szemben a juxtamedulláris nefronok (mintegy 15%) Henle-kacsa rendkívül hosszú, és mélyen behatol a velőállományba, egészen a papilla csúcsáig. Ezek a hosszú kacsok kulcsfontosságúak az ozmotikus gradiens kialakításában és fenntartásában, ami elengedhetetlen a koncentrált vizelet képzéséhez.
A Henle-kacs maga három fő szegmensre osztható, amelyek mindegyike eltérő permeabilitási tulajdonságokkal és transzporterekkel rendelkezik, lehetővé téve a célzott víz- és sómozgást.
A leszálló szár: Vízáteresztő kapu
A Henle-kacs leszálló szára közvetlenül a proximális tubulusból indul, és a velőállomány felé halad. Jellemzője, hogy rendkívül áteresztő a víz számára, köszönhetően a membránjában található aquaporin-1 (AQP1) vízközvetítő csatornáknak. Ezzel szemben viszonylag átjárhatatlan a sók és az urea számára.
Ahogy a szűrlet lefelé áramlik a leszálló szárban, a velőállomány egyre növekvő ozmotikus koncentrációja miatt a víz passzívan kiáramlik a tubulusból az interstitiális térbe. Ez a folyamat a szűrlet koncentrációjának növekedését eredményezi, miközben a térfogata csökken. A szűrlet ozmolaritása a leszálló szár végére elérheti a 1200 mOsm/kg-ot is, ami négyszerese a vérplazma ozmolaritásának.
Ennek a szegmensnek a funkciója tehát alapvető a vízvisszaszívásban, és előkészíti a terepet a következő szakaszban zajló sótranszportnak. Az aquaporin-1 hiánya súlyosan károsíthatja a vese koncentráló képességét, ami jelentős vízháztartási zavarokhoz vezethet.
A vékony felszálló szár: Sók passzív áramlása
A leszálló szár fordulója után következik a vékony felszálló szár, amely felfelé, a kéregállomány felé halad. Ez a szegmens éppen ellenkező tulajdonságokkal rendelkezik, mint a leszálló szár: teljesen átjárhatatlan a víz számára, de bizonyos mértékben átjárható a sók (különösen a nátrium-klorid) és az urea számára.
Mivel a szűrlet rendkívül koncentrált a leszálló szár végén, és a velőállomány interstitiuma is magas ozmolaritású, a nátrium és klorid ionok passzívan diffundálnak ki a tubulusból az interstitiális térbe, követve a koncentrációgradienset. Ez a passzív sóáramlás tovább hozzájárul a velőállomány ozmotikus grádiensének fenntartásához.
A vékony felszálló szár tehát a sók kiválasztásában játszik szerepet, anélkül, hogy vizet veszítene. Ez a mechanizmus kulcsfontosságú a szűrlet hígításában, mielőtt az a vastag felszálló szárba jutna, ahol az aktív transzport veszi át a főszerepet.
A vastag felszálló szár: Az aktív transzport motorja
A Henle-kacs vastag felszálló szára a vékony felszálló szár folytatása, és a kéregállomány felé haladva vastagabbá válik, a sejtekben több mitokondriummal és aktív transzporterrel rendelkezik. Ez a szegmens is teljesen átjárhatatlan a víz számára, még az ADH jelenlétében is.
Azonban a vastag felszálló szár intenzíven és aktívan visszaszívja az ionokat a szűrletből az interstitiális térbe. A legfontosabb transzporter itt a Na+-K+-2Cl- kotranszporter (NKCC2), amely egyidejűleg szállít egy nátrium-, egy kálium- és két kloridiont a tubulus lumenéből a sejtbe.
A Henle-kacs vastag felszálló szára kulcsfontosságú az ozmotikus gradiens kialakításában, hiszen itt történik a nátrium, kálium és klorid ionok aktív visszaszívása a vizeletből, vízveszteség nélkül.
Az NKCC2 működését a Na+/K+-ATPáz táplálja, amely a nátriumot aktívan pumpálja ki a sejtből az interstitiumba, fenntartva a nátrium-grádienst a sejt belsejében. Ez a folyamat hatalmas mennyiségű sót von ki a szűrletből, anélkül, hogy víz követné azt, ami a szűrlet jelentős hígulásához vezet.
Amikor a szűrlet elhagyja a vastag felszálló szárat és a disztális tubulusba jut, ozmolaritása akár 100 mOsm/kg-ra is csökkenhet, ami jelentősen alacsonyabb, mint a vérplazmáé. Emiatt nevezik ezt a szegmenst „hígító szegmensnek” is. Az itt létrehozott híg vizelet elengedhetetlen a szervezet vízháztartásának finomhangolásához.
Az ellenáramú sokszorozó mechanizmus: A vese zsenialitása
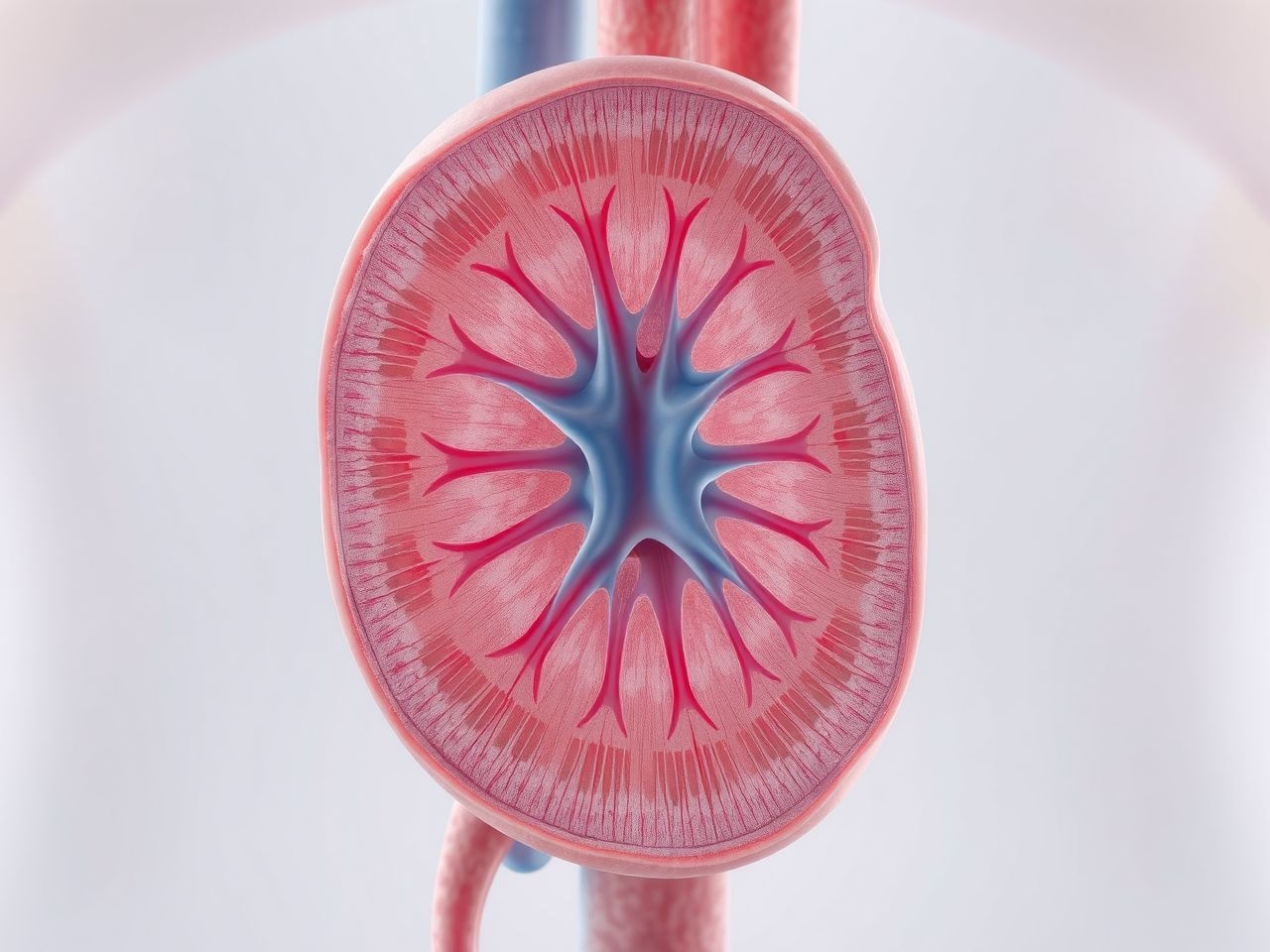
A Henle-kacs egyedülálló, U alakú szerkezete nem véletlen. Ez a forma teszi lehetővé az úgynevezett ellenáramú sokszorozó mechanizmus működését, ami a vese koncentráló képességének alapja. Ez a rendszer egy összetett interakció a Henle-kacs leszálló és felszálló szárai, valamint a vesemedulla interstitiális folyadéka között.
Az ellenáramú sokszorozó rendszer célja, hogy fenntartsa és fokozza a velőállományban lévő ozmotikus grádienst, amely a kéregtől a velőállomány csúcsa felé haladva egyre koncentráltabbá válik. Ez a gradiens nélkülözhetetlen ahhoz, hogy a gyűjtőcsatornákból víz szívódjon vissza, és koncentrált vizelet képződjön az antidiuretikus hormon (ADH) hatására.
Az ozmotikus gradiens kialakulása
A mechanizmus a következőképpen működik:
- A vastag felszálló szár aktívan pumpálja ki a nátrium-kloridot a tubulus lumenéből az interstitiumba. Mivel ez a szegmens vízre átjárhatatlan, a szűrlet hígul, az interstitium pedig koncentrálódik.
- Ez a koncentrált interstitium vonzza a vizet a leszálló szárból, amely vízre áteresztő. Ahogy a víz kiáramlik, a leszálló szárban lévő szűrlet koncentrációja növekszik.
- A leszálló szár alján a szűrlet rendkívül koncentrált. Ahogy ez a koncentrált szűrlet belép a felszálló szárba, a benne lévő magas sókoncentráció lehetővé teszi, hogy még több só diffundáljon vagy pumpálódjon ki, tovább fokozva az interstitium koncentrációját.
Ez a folyamat ciklikusan ismétlődik, és minden egyes ciklusban „sokszorozódik” a koncentrációkülönbség a tubulus és az interstitium között. Ennek eredményeként a velőállomány interstitiumának ozmolaritása a kéregtől a mélyebb velőállomány felé haladva fokozatosan növekszik, elérve akár a 1200 mOsm/kg-ot is a papilla csúcsán.
A vasa recta szerepe: Az ozmotikus gradiens megőrzése
Az ozmotikus gradiens fenntartásában kulcsfontosságú szerepet játszanak a vasa recta, amelyek a juxtamedulláris nefronok Henle-kacsait körülölelő kapillárisok. Ezek a speciális erek szintén U alakú hurkot képeznek, és ellenáramú elrendezésben futnak a Henle-kacsokkal.
A vasa recta rendkívül áteresztő a víz és az oldott anyagok számára. Ahogy a vér lefelé áramlik a vasa recta leszálló ágában a velőállomány egyre koncentráltabb régióiba, sókat vesz fel és vizet veszít. Amikor azonban a vér felfelé áramlik a vasa recta felszálló ágában, a velőállomány kevésbé koncentrált régióiba, sót veszít és vizet vesz fel.
Az ellenáramú sokszorozó rendszer működése nélkülözhetetlen a koncentrált vizelet termeléséhez, lehetővé téve a szervezet számára a vízháztartás precíz szabályozását extrém körülmények között is.
Ez az ellenáramú csere megakadályozza, hogy a vasa recta „kimossa” az ozmotikus grádienst a velőállományból. A vasa recta tehát nem hozza létre az ozmotikus grádienst, hanem megőrzi azt, miközben elegendő vért szállít a velőállomány sejtjeinek oxigén- és tápanyagellátásához.
Az urea (karbamid) újrahasznosításának szerepe
Az ellenáramú sokszorozó mechanizmus hatékonyságát jelentősen fokozza az urea (karbamid) újrahasznosítása. Az urea a fehérjeanyagcsere végterméke, és szintén fontos ozmotikusan aktív anyag a vesemedullában.
Az urea passzívan filtrálódik a glomerulusban, majd jelentős része visszaszívódik a proximális tubulusban. A Henle-kacsban azonban az urea koncentrációja ismét megnő, különösen a leszálló szárban, mivel a víz kiáramlik. A vékony és vastag felszálló szár viszonylag átjárhatatlan az urea számára, így az tovább halad a disztális tubulusba.
A gyűjtőcsatornákban az ADH hatására a víz visszaszívódik, ami tovább koncentrálja az ureát. A gyűjtőcsatorna legvégén, a belső velőállományban található speciális urea transzporterek (UT1 és UT3) lehetővé teszik az urea passzív diffúzióját az interstitiális térbe. Ez az interstitiális urea aztán diffundálhat a Henle-kacs vékony felszálló és leszálló szárába, „újrahasznosítva” magát, és hozzájárulva a velőállomány magas ozmolaritásához.
Az urea újrahasznosításának ez a körforgása kulcsfontosságú az ozmotikus gradiens maximális fenntartásában, különösen vízhiányos állapotokban, amikor a szervezetnek rendkívül koncentrált vizeletet kell termelnie. Ez a mechanizmus mintegy 50%-ban felelős a velőállomány teljes ozmolaritásáért.
A Henle-kacs működésének szabályozása
A Henle-kacs működése nem egy statikus folyamat, hanem dinamikusan szabályozott, hogy a szervezet folyamatosan alkalmazkodni tudjon a változó igényekhez, mint például a folyadékbevitelhez vagy a sófogyasztáshoz. Számos hormonális és lokális mechanizmus befolyásolja a kacs aktivitását.
Az antidiuretikus hormon (ADH) és a vízháztartás
Az antidiuretikus hormon (ADH), más néven vazopresszin, az agyalapi mirigy hátsó lebenyéből szabadul fel, elsősorban a plazma ozmolaritásának növekedésére vagy a vérnyomás csökkenésére válaszul. Bár az ADH elsősorban a gyűjtőcsatornák vízáteresztő képességét szabályozza, hatása szorosan összefügg a Henle-kacs által létrehozott ozmotikus gradienssel.
Az ADH a gyűjtőcsatornák sejtjeiben az aquaporin-2 (AQP2) vízközvetítő csatornák beépülését stimulálja a membránba, így azok vízre áteresztővé válnak. Ennek eredményeként a víz passzívan kiáramlik a gyűjtőcsatornákból a Henle-kacs által fenntartott magas ozmolaritású velőállományi interstitiumba, ami koncentrált vizelet képződését eredményezi.
Ha a Henle-kacs nem lenne képes létrehozni ezt az ozmotikus grádienst, az ADH hiába tenné áteresztővé a gyűjtőcsatornákat, a víz nem tudna visszaszívódni, és a szervezet nagy mennyiségű híg vizeletet ürítene, ami dehidratációhoz vezetne. Az ADH tehát a Henle-kacs munkájára építve fejti ki hatását.
A renin-angiotenzin-aldoszteron rendszer (RAAS) és a Henle-kacs
A renin-angiotenzin-aldoszteron rendszer (RAAS) egy komplex hormonális kaszkád, amely kulcsszerepet játszik a vérnyomás és a folyadék-elektrolit egyensúly szabályozásában. Bár az aldoszteron elsősorban a disztális tubulusra és a gyűjtőcsatornákra hat, a RAAS aktiválódása közvetett módon befolyásolja a Henle-kacs működését is.
Az angiotenzin II, a RAAS egyik aktív komponense, növelheti a proximális tubulusban a nátrium visszaszívását, ami befolyásolhatja a Henle-kacsba érkező szűrlet összetételét. Ezenkívül az angiotenzin II befolyásolhatja a medulláris véráramlást a vasa recta-ban, ami hatással van az ozmotikus gradiens fenntartására.
Az aldoszteron hatására a disztális tubulusban és a gyűjtőcsatornákban fokozódik a nátrium visszaszívása és a kálium szekréciója. Bár ez közvetlenül nem a Henle-kacsban történik, a RAAS-on keresztül történő volumen- és elektrolit-szabályozás végső soron befolyásolja a Henle-kacs által fenntartott ozmotikus gradiens jelentőségét és a vizelet végső koncentrációját.
Tubuloglomeruláris visszacsatolás (TGF): A lokális kontroll
A tubuloglomeruláris visszacsatolás (TGF) egy lokális szabályozó mechanizmus, amely a Henle-kacs vastag felszálló szárának elején található macula densa sejtek segítségével működik. A macula densa sejtek érzékelik a szűrlet nátrium-klorid koncentrációját a tubulus lumenében.
Ha a glomerulus filtrációs rátája (GFR) megnő, a szűrlet gyorsabban halad át a proximális tubuluson, és kevesebb nátrium-klorid szívódik vissza. Ennek eredményeként a macula densához magasabb nátrium-klorid koncentrációjú szűrlet érkezik. A macula densa sejtek ezt érzékelve vazokonstriktor anyagokat (pl. adenozin) bocsátanak ki, amelyek összehúzzák az afferens arteriolát (az artériát, amely a glomerulusba vezeti a vért).
Az afferens arteriola szűkülete csökkenti a glomerulusba jutó véráramlást és a glomerulus filtrációs rátáját, ezzel normalizálva a macula densához érkező nátrium-klorid koncentrációját. Ez a negatív visszacsatolási hurok biztosítja, hogy a GFR viszonylag stabil maradjon, és optimalizálja a Henle-kacs terhelését, megakadályozva a túlzott só- és vízveszteséget.
Klinikai relevanciája és patológiás állapotai
A Henle-kacs pótolhatatlan szerepe a vízháztartás és az elektrolit-egyensúly fenntartásában azt jelenti, hogy bármilyen diszfunkciója súlyos klinikai következményekkel járhat. Számos gyógyszer és betegség befolyásolja közvetlenül vagy közvetve a Henle-kacs működését.
Kacsdiuretikumok: A Henle-kacs gátlása gyógyszerekkel
A kacsdiuretikumok a legpotensebb vízhajtó gyógyszerek közé tartoznak, és széles körben alkalmazzák őket olyan állapotok kezelésére, mint az ödéma (szív-, vese- vagy májelégtelenség miatt), a magas vérnyomás és a hyperkalcemia. Hatásmechanizmusuk a Henle-kacs vastag felszálló szárának NKCC2 kotranszporterének gátlásán alapul.
A kacsdiuretikumok, mint például a furoszemid, a Henle-kacs vastag felszálló szárában található NKCC2 transzporter blokkolásával fejtik ki hatásukat, jelentősen növelve a vizeletürítést és csökkentve az ödémát.
Az NKCC2 gátlása megakadályozza a nátrium, kálium és klorid ionok aktív visszaszívását, ami megnöveli az oldott anyagok koncentrációját a tubulus lumenében. Mivel a vastag felszálló szár vízre átjárhatatlan, ez a sóveszteség megakadályozza az ozmotikus gradiens kialakulását vagy fenntartását a vesemedullában.
Ennek következtében a gyűjtőcsatornákból sem tud visszaszívódni a víz, még az ADH jelenlétében sem, ami hatalmas mennyiségű híg vizelet ürítéséhez vezet. A kacsdiuretikumok mellékhatásai közé tartozhat a hipokalémia (alacsony káliumszint), hiponatremia (alacsony nátriumszint) és a metabolikus alkalózis.
Bartter szindróma: A Henle-kacs örökletes rendellenessége
A Bartter szindróma egy ritka, örökletes betegség, amelyet az NKCC2 kotranszporter vagy a kapcsolódó ioncsatornák (pl. káliumcsatornák) genetikai hibája okoz a Henle-kacs vastag felszálló szárában. Ez a hiba lényegében „belső kacsdiuretikum-hatást” eredményez.
A betegségben szenvedők képtelenek hatékonyan visszaszívni a sót a vastag felszálló szárban, ami krónikus só- és vízháztartási zavarokhoz vezet. Jellemző tünetei a súlyos sóvesztés, nagy mennyiségű híg vizelet ürítése (polyuria), szomjúság (polydipsia), alacsony káliumszint (hypokalemia), metabolikus alkalózis és gyakran alacsony vérnyomás.
A Bartter szindróma súlyos fejlődési rendellenességeket és növekedési elmaradást okozhat gyermekeknél. A kezelés a só- és káliumpótlásra, valamint a prosztaglandin szintézis gátlására irányul, amely enyhítheti a renin-angiotenzin-aldoszteron rendszer túlműködését.
Diabetes insipidus: A Henle-kacs közvetett érintettsége
A diabetes insipidus egy olyan állapot, amelyet az ADH hiánya (centrális diabetes insipidus) vagy a vese ADH-ra való érzéketlensége (nefrogén diabetes insipidus) okoz. Bár a probléma elsősorban az ADH-val és a gyűjtőcsatornákkal kapcsolatos, a Henle-kacs által létrehozott ozmotikus gradiens nélkülözhetetlen szerepe miatt fontos megemlíteni.
A diabetes insipidusban szenvedő betegek nagy mennyiségű, rendkívül híg vizeletet ürítenek, mert a gyűjtőcsatornákból nem tud visszaszívódni a víz. Ez súlyos dehidratációhoz és elektrolit-zavarokhoz vezethet. A Henle-kacs maga általában normálisan működik, létrehozva a szükséges ozmotikus grádienst.
A probléma abban rejlik, hogy az ADH hiánya vagy a gyűjtőcsatornák ADH-ra való válaszának hiánya miatt a Henle-kacs által létrehozott gradiens nem hasznosul. A Henle-kacs tehát megteremti a feltételeket a vízvisszaszívásra, de az utolsó lépés, a gyűjtőcsatornákban történő vízvisszaszívás akadályozott.
Veseelégtelenség és a Henle-kacs diszfunkciója
A veseelégtelenség, legyen az akut vagy krónikus, számos okból kifolyólag érintheti a Henle-kacs működését. A veseszövet károsodása, különösen a velőállományban, közvetlenül ronthatja az ozmotikus gradiens fenntartásának képességét. Ez a vese koncentráló képességének csökkenéséhez vezet, ami polyuriát és elektrolit-zavarokat okoz.
Például akut tubuláris nekrózis (ATN) esetén, amely gyakran ischaemia vagy toxikus anyagok hatására alakul ki, a Henle-kacs sejtjei károsodhatnak, és az NKCC2 transzporter funkciója romolhat. Ez súlyos folyadék- és elektrolit-egyensúlyi zavarokhoz vezet, és hozzájárul a veseelégtelenség patofiziológiájához.
Krónikus veseelégtelenségben a nephronok számának csökkenése és a megmaradt nephronok hiperfiltrációja is befolyásolhatja a Henle-kacs működését. A fennmaradó nephronok Henle-kacsai túlterhelődhetnek, ami idővel kimerülhet, és tovább ronthatja a vese koncentráló képességét. A Henle-kacs tehát kulcsfontosságú célpont és indikátor a vesebetegségek progressziójában.
A Henle-kacs evolúciós jelentősége
A Henle-kacs nem csak anatómiai csoda, hanem az evolúció egyik legkiemelkedőbb adaptációja is a szárazföldi élethez. A vízháztartás fenntartása a szárazföldi környezetben az egyik legnagyobb kihívás az állatok számára, és a Henle-kacs kifejlődése döntő lépés volt ebben a folyamatban.
Azok az állatok, amelyek sekély vízben vagy tengerben élnek, és nem szembesülnek jelentős vízhiánnyal (pl. halak), nem rendelkeznek Henle-kaccsal. Ehelyett egyszerűbb nefronjaik vannak, amelyek nagy mennyiségű híg vizeletet termelnek. A kétéltűek Henle-kacsa is viszonylag rövid és egyszerű.
Azonban a hüllők, madarak és emlősök, amelyek szárazföldi környezetben élnek, és hatékonyan kell vizet takarékoskodniuk, mind rendelkeznek Henle-kaccsal. Minél szárazabb a környezet, annál hosszabb és fejlettebb a Henle-kacs, különösen a juxtamedulláris nefronokban.
Például a sivatagi emlősök, mint a kenguru patkány, rendkívül hosszú Henle-kacsokkal rendelkeznek, amelyek mélyen benyúlnak a vesemedullába, lehetővé téve számukra, hogy rendkívül koncentrált vizeletet termeljenek, és minimálisra csökkentsék a vízveszteséget. Ez az adaptáció kulcsfontosságú a túlélésükhöz a vízhiányos környezetben.
Ez az evolúciós nyomás tette a Henle-kacsot a vízháztartás szabályozásának mesterművévé, amely lehetővé teszi a szárazföldi gerincesek számára, hogy fennmaradjanak és virágozzanak változatos és gyakran kihívást jelentő környezetekben.
Összefüggések a nefron más részeivel
Bár a Henle-kacs önmagában is rendkívül fontos, működése elválaszthatatlanul összefügg a nefron többi részének tevékenységével. A vizeletképzés egy integrált folyamat, ahol minden szegmens hozzájárul a végső eredményhez.
A proximális tubulus és a Henle-kacs
A proximális tubulus a Henle-kacs „előszobája”. Itt történik a primer szűrlet térfogatának és összetételének jelentős módosítása. Körülbelül a szűrlet 65-70%-a, valamint szinte az összes glükóz és aminosav visszaszívódik a proximális tubulusban.
Ez a masszív visszaszívás biztosítja, hogy a Henle-kacsba érkező szűrlet már egy koncentráltabb, de még mindig izoozmotikus folyadék legyen. Ha a proximális tubulus diszfunkcionális lenne, a Henle-kacsra sokkal nagyobb terhelés hárulna, ami felborítaná a vese finom egyensúlyát.
A proximális tubulusban történő só- és vízvisszaszívás mértéke közvetlenül befolyásolja a Henle-kacsba jutó szűrlet mennyiségét és ozmolaritását, ami alapvető a Henle-kacs hatékony működéséhez és az ellenáramú sokszorozó mechanizmus fenntartásához.
A disztális tubulus és a Henle-kacs
A Henle-kacs vastag felszálló szárából kilépő szűrlet rendkívül híg, ozmolaritása akár 100 mOsm/kg is lehet. Ez a híg szűrlet jut a disztális tubulusba, ahol a „finomhangolás” történik.
A disztális tubulusban további nátrium-klorid visszaszívás zajlik, amelyet a tiaciddiuretikumok gátolnak. Itt történik a kalcium visszaszívásának szabályozása is, a parathormon (PTH) hatására. A disztális tubulus tehát tovább módosítja a szűrlet összetételét, mielőtt az a gyűjtőcsatornákba kerülne.
A Henle-kacs által létrehozott híg szűrlet elengedhetetlen ahhoz, hogy a disztális tubulus és a gyűjtőcsatornák hatékonyan tudjanak reagálni a hormonális jelekre, és pontosan szabályozzák a végső vizelet ozmolaritását. A Henle-kacs nélkül a disztális tubulus nem tudná elérni a kívánt vizeletkoncentrációt.
A gyűjtőcsatornák és a Henle-kacs
A gyűjtőcsatornák jelentik a vizeletképzés utolsó szakaszát, és itt dől el a vizelet végső koncentrációja. A Henle-kacs által létrehozott ozmotikus gradiens a vesemedullában kulcsfontosságú a gyűjtőcsatornák működéséhez.
Az ADH jelenlétében a gyűjtőcsatornák áteresztővé válnak a víz számára, és a víz passzívan kiáramlik a csatornákból a Henle-kacs által fenntartott magas ozmolaritású interstitiumba. Minél nagyobb az ozmotikus gradiens, annál több víz szívódhat vissza, és annál koncentráltabb lesz a vizelet.
A gyűjtőcsatornákban történő urea újrahasznosítás is szorosan összefügg a Henle-kacs működésével, hiszen az urea hozzájárul az ozmotikus gradienshez, amelyre a gyűjtőcsatornák támaszkodnak. Ez a szinergikus kapcsolat teszi lehetővé a szervezet számára, hogy rendkívül széles tartományban szabályozza a vizelet koncentrációját, a híg vizelettől a maximálisan koncentráltig.
Kutatási perspektívák és jövőbeli irányok
A Henle-kacs működésének és jelentőségének kutatása továbbra is aktív terület a nefrológiában és a fiziológiában. Bár alapvető mechanizmusait már régóta ismerjük, a molekuláris szintű részletek, a szabályozási hálózatok és a patológiás állapotok finomabb aspektusai még mindig tartogatnak felfedeznivalót.
A genetikai kutatások például továbbra is új géneket és mutációkat azonosítanak, amelyek a Henle-kacs transzportereinek vagy ioncsatornáinak működését befolyásolják. Ezek a felfedezések mélyebb betekintést nyújtanak olyan betegségek etiológiájába, mint a Bartter szindróma, és új célpontokat kínálhatnak a célzott terápiák fejlesztéséhez.
A gyógyszerfejlesztés területén is folyamatosan keresnek új vegyületeket, amelyek specifikusabban vagy kevesebb mellékhatással befolyásolják a Henle-kacs funkcióját. Például az NKCC2 transzporter szelektívebb gátlói, vagy olyan szerek, amelyek modulálják a vízközvetítő csatornák aktivitását, ígéretes terápiás lehetőségeket jelenthetnek.
A képalkotó eljárások fejlődése is hozzájárul a Henle-kacs in vivo vizsgálatához. Új MRI technikák és funkcionális képalkotó módszerek lehetővé teszik a vese velőállományának ozmotikus gradiensének és a véráramlásnak a non-invazív mérését, ami segíthet a Henle-kacs diszfunkciójának korai felismerésében.
| Kutatási terület | Lehetséges áttörések |
|---|---|
| Genetikai betegségek | Célzott terápiák Bartter és Gitelman szindrómára |
| Transzporter mechanizmusok | Új diuretikumok fejlesztése, mellékhatások csökkentése |
| Veseelégtelenség megelőzése | A Henle-kacs diszfunkciójának korai felismerése és kezelése |
| Regeneratív medicina | Mesterséges nefronok vagy Henle-kacs szegmensek létrehozása |
| Molekuláris farmakológia | A Henle-kacs transzportereinek finomhangolása a betegségek kezelésére |
A regeneratív medicina és a veseszövet-mérnökség területén is zajlanak kutatások, amelyek célja mesterséges nefronok vagy veseszegmensek létrehozása, amelyek magukban foglalják a funkcionális Henle-kacsot. Ez a jövőben áttörést hozhat a veseelégtelenség kezelésében és a dialízis alternatívájának megteremtésében.
A Henle-kacs tehát nem csupán egy anatómiai struktúra, hanem egy dinamikus, szabályozott és rendkívül adaptív rendszer, amelynek megértése alapvető a vese egészségének és a szervezet folyadékháztartásának fenntartásához. A folyamatos kutatás és a tudományos fejlődés reményt ad arra, hogy a jövőben még hatékonyabban tudjuk kezelni a Henle-kacs diszfunkciójával járó betegségeket.







